Immunoglobulin A (IgA) comprises 5 to 15% of the serum immunoglobulins and has a half-life of 6 days.
It has a molecular weight of 160 kDa and a basic four-chain monomeric structure (two L and two H) chain proteins.
However, it can occur as monomers, dimers, trimers, and polymers.
It contains a heavy chains and ? or ? light chains.
Immunoglobulin alpha (a) chain is a 58-kDa molecular weight with 470-amino acid residue heavy polypeptide chain that confers class specificity on immunoglobulin A molecules.
The heavy chain is divisible into three constant domains, designated CH1, CH2, and CH3, and one variable domain, designated VH.
The hinge region is the area of the H chains between CH1 and CH2 domains and is held together by disulfide bonds.
Each L chain consists of one variable domain, VL, and one constant domain, CL.
The monomeric structural unit of IgA comprises two identical heavy chains and two identical light chains arranged into two Fab regions and an Fc region, separated by a flexible hinge region.
The paired variable regions at the tips of the Fab arms are responsible for antigen recognition, while the Fc region mediates interaction with various receptors and effector molecules.
The heavy and light chains fold up into globular domains, four in each heavy chain (from the N-terminus VH, Ca1, Ca2, and Ca3) and two in each light chain (VL and CL), each with a characteristic tertiary structure of anti-parallel ß sheets termed the immunoglobulin fold.
The majority of IgA in the secretions is polymeric, mainly in the form of dimers.
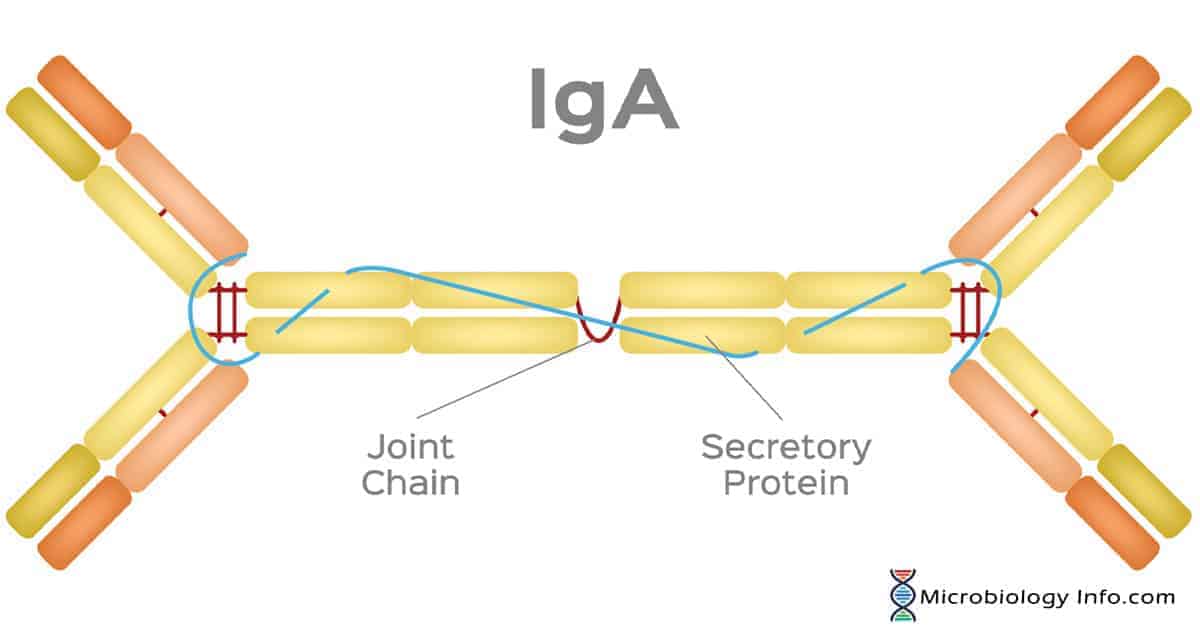
The CH3 domains of IgA have short tailpieces to which the J-chain binds via disulfide bonds whereas the secretory component is disulfide bonded to one of the CH2 domains of the dimer.
In addition to the ? or ? L-chains and the IgA heavy chain alpha, IgA also contains two other polypeptide chains including a 15KD joining chain (J chain) and a 70KD secretory component chain produced in epithelial cells and involved in the transcellular transport of Secretory IgA.
When IgA is found in secretions it also has protein associated with it called the secretory piece or T piece.
S-IgA is secretory component (SC) is an additional component of IgA which is a polypeptide of 75- 80 kDa comprising the extracellular proteolytic fragment of the receptor responsible for transport of dimeric IgA into the secretions, namely the polymeric immunoglobulin receptor (pIgR).
S-IgA is sometimes referred to as 11S immunoglobulin.
The two four-chain units are held together by the J-chain through disulfide bridges.
SC and J chain are both disulphide-linked to the Fc region of the IgA, although not to one another.
Secretory component facilitates IgA transport across epithelial cells (transcytosis) and protects secretory IgA released into the lumen of the gut from proteolytic digestion by enzymes in the secretions.
Properties of Immunoglobulin A (IgA)
Immunoglobulin A (IgA) is the most abundant immunoglobulin found in humans.
IgA, at concentrations of about 2–4 mg/ml, is considered as the second most prevalent antibody in serum after Immunoglobulin G (IgG), which is normally present at about 12 mg/ml.
IgA constitutes 10%–15% of the total immunoglobulin in serum and is the predominant immunoglobulin class found in external secretions which plays key roles in immune protection.
In the resistance against infection , Immunoglobulin A (IgA) is the first line of defense. It works via inhibiting bacterial and viral adhesion to epithelial cells and by neutralization of viruses and bacterial toxins, both extra- and intracellularly.
IgA also helps to eliminate pathogens or antigens via an IgA-mediated excretory pathway where binding to IgA is followed by poly immunoglobulin receptor-mediated transport of immune complexes.
IgA exists in two forms, monomeric and polymeric.
Serum IgA is predominantly monomeric IgA1 which is produced in the bone marrow, while in external secretions most of the locally produced IgA is polymeric with a relative increase in the proportion of IgA2 comprising mainly dimeric forms connected by a joining peptide chain.
Secretory IgA exists in a dimer form and provides the primary defense mechanism against some local infections because of its abundance in mucosal secretions (e.g., saliva and tears).
IgA appears selectively in the seromucous secretions, such as saliva, tears, nasal fluids, sweat, colostrum, milk, and secretions of the lung, genitourinary and gastrointestinal tracts, where it defends the exposed external surfaces of the body against attack by microorganisms.
This is an important function of IgA as approximately 40 mg of secretory IgA/kg body weight is transported daily through the human intestinal crypt epithelium to the mucosal surface as compared with a total daily production of IgG of 30 mg/kg.
The deficiency of IgA in individuals has an increased incidence of respiratory infections associated with a lack of secretory IgA in the respiratory system.
Subclasses
In humans, two subclasses of IgA, termed IgA1 and IgA2, exist, each the product of a separate gene reflecting two separate a chain isotypes.
They differ in the molecular mass of the heavy chains and in their concentration in serum.
A major difference between the subclasses lies in the hinge region, which is greatly extended in IgA1.
Numerous sequence differences are found in their heavy chain constant regions.
Their heavy chain constant region sequences differ at a number of points along their length.
IgA1
IgA1 comprises approximately 85% of total IgA concentration in serum.
IgA1 molecules are marked out by the length of their hinge regions, flexible stretches of polypeptide at the antibody’s core that separate the regions responsible for antigen binding and effector capability.
This elongated hinge region increases the sensitivity of IgA1 to bacterial proteases in spite of partial protection by glycans. Such increased protection against protease digestion may explain why IgA2 predominates in the many mucosal secretions, such as the genital tract, whereas more than 90% of serum IgA is in the form of IgA1.
Although IgA1 shows a broad resistance against several proteases, there are some that can affect/splice on the hinge region.
IgA1 shows a good immune response to protein antigens and, to a lesser degree, to lipopolysaccharides and polysaccharides.
IgA2
IgA2 comprises 15% of total IgA in serum and plays a crucial role in the mucosa of the eyes, airways and the gastrointestinal tract to fight against polysaccharide and lipopolysaccharide antigens.
It also shows good resistance to proteolysis and many bacterial proteases, supporting the importance of IgA2 in fighting bacterial infections.
In addition, the IgA2 has two allotypic variants designated IgA2m(1) and IgA2m(2) that have been defined serologically and at the molecular level are widely distributed in the human gene pool.
The most notable difference relates to the fact that while IgA2m(2) has the disulphide bridges linking light and heavy chains typical of most immunoglobulins, these are generally lacking in IgA2m(1).
IgA synthesis and distribution
The IgA is synthesized locally by plasma cells and get dimerized intracellularly together with a cysteine – rich polypeptide called J chain.
Dimeric IgA binds strongly to a receptor for polymeric Ig present in the membrane of mucosal epithelial cells.
The complex is then actively endocytosed, transported across the cytoplasm and secreted into the external body fluids after cleavage of the pIgR peptide chain.
The fragment of the receptor remaining bound to the IgA is termed as secretory component and the whole molecule is called secretory IgA.
Secretory component is synthesized in epithelial and glandular cells, in which it is first expressed as a basolateral membrane receptor for J-chain-containing polymeric immunoglobulins, and has therefore been referred to as “polymeric immunoglobulin receptor.”
During transepithelial transport in endocytic vesicles, polymeric IgA usually becomes covalently linked through formation of disulfide bonds to the secretory component, which is proteolytically cleaved from its transmembrane segment before exocytotic release of the complete secretory IgA at the apical surface.
The daily production of secretory IgA is greater than that of any other immunoglobulin class.
Every day, a human secretes from 5 g to 15 g of secretory IgA into mucous secretions.
The plasma cells that produce IgA preferentially migrate to subepithelial tissue, where the secreted IgA binds tightly to a receptor for polymeric immunoglobulin molecules.
IgA is the product of local synthesis at the mucosal surfaces, which serve as the main source of antigenic material for the body.
The mucosal immune system such as the gut-associated lymphoid tissue (GALT) is highly specialized and functions largely independently of the systemic immune system.
Serum IgA is predominantly (~90%) monomeric IgA1 that is produced in the bone marrow, while in external secretions most of the locally produced IgA is polymeric with a relative increase in the proportion of IgA2 types.
The lymphocytes that produce monomeric or polymeric IgA of IgA1 or IgA2 subclass are characteristically distributed in various lymphoid and non-lymphoid tissues.
The differential interaction of monomeric and polymeric IgA molecules with various cells leads to their selective distribution in body fluids and possibly to differences in their effector functions.
Secretory and serum IgA are therefore molecules with different biochemical and immunochemical properties produced by cells with different organ distributions.
IgA production is regulated by T cells. T cell derived transforming growth factor ß (TGFß) has been identified as the primary cytokine that promotes B cells to switch from IgM bearing cells to IgA bearing cells.
Functions of Immunoglobulin A (IgA)
Secretory IgA serves an important effector function at mucous membrane surfaces, which are the main entry sites. It primarily protects mucous membranes as IgA can cross the epithelial layer and enter into body secretion providing local immunity.
Secretory IgA provides the primary defense mechanism against some local infections because of its abundance in mucosal secretions (e.g., saliva and tears).
Secretory IgA has been shown to provide an important line of defense against bacteria such as Salmonella, Vibrio cholerae, and Neisseria gonorrhoeae and viruses such as polio, influenza, and reovirus.
Secretory IgA can also inhibit inflammatory effects of other immunoglobulins.
It prevents the passage of foreign substances into the circulatory system of the body.
In body secretion IgA neutralizes viruses and prevents attachment on the host surface.
The very high concentration of secretory IgA in human colostrum and milk suggests strongly that it must play an important role in the passive (and possibly active) immune protection of the newborn. Mothers’ milk contains S-IgA against a wide variety of microbial antigens and these antibodies are able to neutralize toxins and viruses.
IgA is resistant to digestion and although a poor activator, can activate the complement pathway when aggregated. IgA does not activate the classical pathway and its role in activation of the alternative pathway remains controversial. However, recently it has become apparent that polymeric IgA (S-IgA) has been shown to activate the lectin pathway of complement activation, by binding the mannose-binding lectin.
Although complement fixation process by IgA does not appear to be a major effector mechanism at the mucosal surface level, the IgA receptor is expressed on neutrophils which may be activated to mediate ADCC locally.
References
Abbas A.K and Lichtman A.H. Cellular and molecular immunology. Fifth edition. Page no.35-43.
Woof J.M and Kerr M.A. (2006). The function of immunoglobulin A in immunity. doi:10.1002/path.1877
Goldsby, R.A., Kindt, T.J., Osborne, B.A. and Kuby J. Kuby Immunology. Fifth edition. Page no. 90-92.
Owen, J. A., Punt, J., & Stranford, S. A and Jones, P.P (2013). Kuby Immunology (7 ed.). New York: W.H. Freeman and Company.
Kerr M.A. (1990). The structure and function of human IgA. Biochem J ;271(2):285-96.
Delves, P.J., Martin, S.J., Burton, D.R. and Roitt, I.M. (2011). Roitt’s Essentail Immunology. 12th edition. A John Wiley and Son’s, Ltd, Publication.Page no: 53-68.
Cruse, J.M. and Lewis, R.E. (2010). Atlas of Immunology. Third edition. CRC Press. Taylor and Francis Group, 6000 Broken Sound Parkway NW, Suite 300, Boca Raton, FL 33487-2742. Page no: 252-306.
Schroeder, H. W., & Cavacini, L. (2010). Structure and function of immunoglobulins. The Journal of allergy and clinical immunology, 125(2 Suppl 2), S41-52.