The immunoglobulin M (IgM) is the largest antibody characterized by the µ heavy chain.
Immunoglobulin M is the third most common serum Ig and exists in two forms: a pentamer comprising approximately 10% of normal human serum Ig content and a membrane-bound monomer expressed on the surface of B lymphocytes for antigen recognition.
The immature B cell called a pre-B cell, expresses only mIgM; later in maturation, mIgD appears and is coexpressed with IgM on the surface of mature B cells before they have been activated by antigen.
IgM is the first immunoglobulin developed during human fetal development at 20 weeks and is restricted from crossing the placenta due to its size. It is synthesized by the neonate and plays a role in the pathogenesis of some autoimmune diseases.
IgM constitutes 5-10% of serum immunoglobulins which accounts for 0.5 to 2 mg/ml serum concentration and has a half-life of 5 to 6 days.
Serum IgM predominates in primary immune responses to most antigens and is a strong complement activator and agglutinator due to its pentameric structure and binds fragment crystallization (Fc) receptors.
IgM is more efficient than IgG in activating classical pathway of complement.
As it can bind to specific antigens without prior immunization, IgM is considered a “natural antibody”. This ability of IgM is likely due its avidity and polyreactivity.
Immunoglobulin M is the antibody formed during the initial response to an antigen or microbe. Following stimulation of antigen, IgM-producing plasma cells migrate from the lymph node to bone marrow, where they turn into long-lived plasma cells. After a certain time period (5 to 10 days), IgM antibodies appear in blood, and peak IgM levels occur at 21 days. During the response, memory B cells are also produced, and they remain in the lymph node germinal centers or recirculate between the lymph node and the spleen.
IgM is predominantly found in the lymph fluid and blood and is a very effective neutralizing agent in the early stages of disease.
Clinically low IgM antibody levels, from undetectable to measuring 20-40 mg/dL, result in persistent, recurrent infection cases.
Structure of Immunoglobulin M
IgM is a glycoprotein of 970KDa molecular weight comprising of basic heavy and light-chain antibody structure but exists in pentameric (secretory) and monomeric configurations.
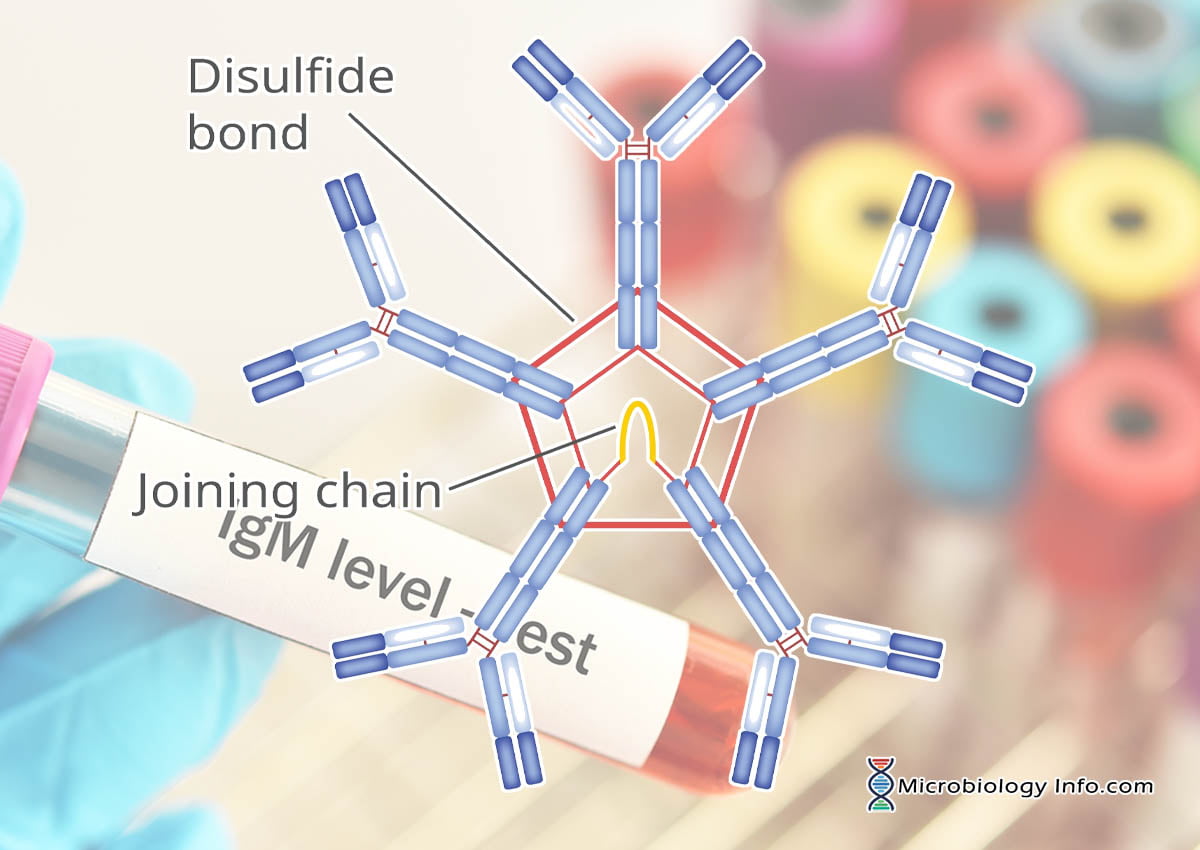
The IgM polymer is composed of subunits in which two heavy chains (µ) are covalently paired with two light chains (L) and a valency of 10
In addition to the disulfide bonds linking the chains together, there are intrachain disulfide links that divide each chain into areas called domains.
The light chains have a molecular weight of 23kDa and have two domains, one variable and one constant.
The µ heavy chains have a molecular weight of approximately 72 kDa and they contain about 57 amino acid residues arranged in five domains and have more oligosaccharide branches and a higher carbohydrate content.
The Fc part of IgM is composed of three Ig domains (Cμ2, Cμ3, and Cμ4) and an additional C-terminal tail piece (tp).
In the pentameric configuration, five antibody units are covalently linked by disulfide bonds at adjacent CH3 domains.
The five-unit IgM molecule has 10 antigen-combining sites that bind antigens with high avidity.
An additional molecule, called a J-chain, a small protein that is involved in IgM assembly and secretion which is attached to the penultimate cysteines of the mu (μ) heavy chains.
The J chain serves two important functions.
Along with the Fc portion of the antibody, it is required for binding to the poly-Ig receptors (pIgRs) that mediate transcellular transport, also called transcytosis, through epithelial cells.
The addition of the J chain causes a conformational change in the molecule, which allows the addition of an 83,000-MW fragment of the pIgR as it traverses through epithelial cells into extracellular fluids. The pIgR fragment, now called the secretory piece, contributes in protecting the secretory IgM (sIgM) from enzymatic degradation.
Upon assembly, the heavy chains are covalently linked in the Cμ2, Cμ3, and Cμ4 (tp) domains by interchain disulfide bridges.
Monomeric IgM is localized on the surfaces of B cell membranes and functions as a B cell antigen receptor (BCR).
The monomeric membrane form of IgM has an additional 41 amino acids in a CH4 domain, and 25 hydrophobic amino acids are found in the transmembrane portion.
The remaining cytoplasmic portion contains 16 polar amino acids.
Functions of Immunoglobulin M
Immunoglobulin M (IgM), the largest pentameric immunoglobulin is used as a marker of recent primary infection for some diseases and can be produced in T independent manner which also aids in disease diagnosis. IgM ELISA test can be performed to detect such antibodies in the serum of patients.
During infection, innate immunity is provided by poly-reactive IgM antibody generated by B cells. IgM antibody acts by quickly recognizing and then initiating an immune response by directly neutralizing pathogens or clearing novel antigens.
Besides mediating the response to immunological challenges, IgM antibody also participates in homeostatic functions of healthy tissue.
When the cells in the body get a natural turnover, the IgM antibody clears apoptotic debris and contributes to inducing the release of IL-10 by the marginal zone B cells, an anti-inflammatory cytokine. In this manner, IgM antibody serves as a mediator of inflammation.
Apoptotic cells release many self-antigens and IgM antibody clearance prevents self-antigen concentrations which could induce an immune response. IgM antibody also contributes to the clearance process by binding to misfolded proteins and altered cells, clearing them via dendritic cells, B cells, and macrophages.
The three important components of the IgM antibody-mediated immune response are activation of complement, recruitment of phagocytic cells, and opsonization. It recognizes a large variety of pathogenic antigens and is highly active in cytotoxic and cytolytic reactions due to its major activation of the complement system.
IgM antibody also acts as an educator of the immune system by transporting antigens to lymph tissues where memory is induced.
The presence of IgM in the fetus or newborn indicates intrauterine infection and its detection is useful in the diagnosis of congenital infections such as Toxoplasmosis, syphilis, rubella, cytomegalovirus infection and HIV infection.
The presence of the J chain in the structure allows IgM to bind to receptors on secretory cells, which transport it across epithelial linings to enter the external secretions that bathe mucosal surfaces. Although IgA is the major isotype found in these secretions, IgM plays an important accessory role as a secretory immunoglobulin.
References
Giraldo, M., Portela, R. W., Snege, M., Leser, P. G., Camargo, M. E., Mineo, J. R., & Gazzinelli, R.T.(2002). Immunoglobulin M (IgM)-glycoinositolphospholipid enzyme-linked immunosorbent assay: an immunoenzymatic assay for discrimination between patients with acute toxoplasmosis and those with persistent parasite-specific IgM antibodies. Journal of clinical microbiology, 40(4), 1400-5.
Müller, R., Gräwert, M. A., Kern, T., Madl, T., Peschek, J., Sattler, M., Groll, M., Buchner, J. (2013). High-resolution structures of the IgM Fc domains reveal principles of its hexamer formation. Proceedings of the National Academy of Sciences of the United States of America, 110(25), 10183-8.
https://www.genscript.com/IgM-antibody.html
Abbas A.K and Lichtman A.H. Cellular and molecular immunology. Fifth edition. Page no.34-43.
Owen, J. A., Punt, J., & Stranford, S. A and Jones, P.P (2013). Kuby Immunology (7 ed.). New York: W.H. Freeman and Company.
Delves, P.J., Martin, S.J., Burton, D.R. and Roitt, I.M. (2011). Roitt’s Essentail Immunology. 12th edition. A John Wiley and Son’s, Ltd, Publication.
Cruse, J.M. and Lewis, R.E. (2010). Atlas of Immunology. Third edition. CRC Press. Taylor and Francis Group, 6000 Broken Sound Parkway NW, Suite 300, Boca Raton, FL 33487-2742.